



Have you ever marvelled at the miraculous feat at which the human body achieves such great complexity. Why do different parts of the eye form specific functions? What tells the retina to be the retina and not the cornea? What is the driving mechanism behind this?Cells may grow and divide in two ways.
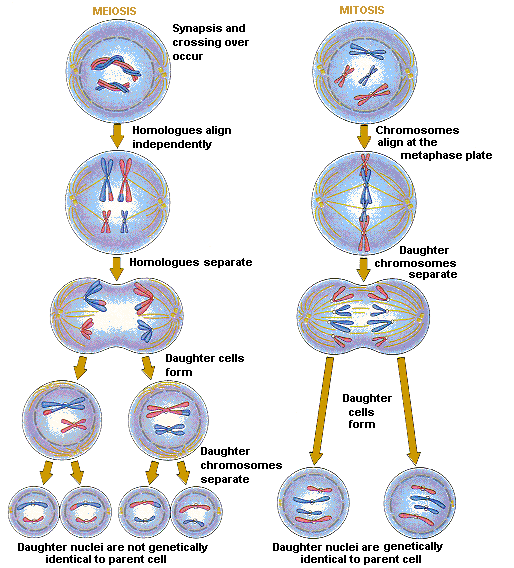
Sex cells divide meiotically whilst somatic cells divide mitotically. Meiotic division is a reductional form of division whereby half of the number of chromosomes are produced giving haploid cells with 23 chromosomes. Thus when two gametes fuse, sperm cell and egg cell, they form a zygote with 46 chromosomes, the original number in the parent cell. Mitotic division however, produces diploid cells which are a clone of the original parent cell with the same amount of chromosomes.In order for cells to develop differently from each other and differentiate into a specific type of cell for a specific function there must be some communication between cells. In the developing embryo, cells become different from their neighbours by two strategies:
- They are induced by their neighbours in a process called induction.
- Daughter cells are distinctly different by asymmetric cell division.

Induction involves a signal from one group of cells which acts upon an adjacent group of cells affecting their development. Induction may be permissive or instructive. Permissive induction occurs when a cell makes only one kind of response to a signal when a sufficient level of signal is reached. An instructive signal however, involves different responses to a signal depending on the concentration of the signal .
The three main ways in which signals may be passed between cells are; secreted diffusible molecules, direct surface molecule interaction and through gap junctions in a direct cell-to-cell contact.
In order to induce adjacent cells to adopt a different cell fate , the signal must alter gene expression in the nucleus and this is called signal transduction. The ability of a cell to be induced also depends on the competence of the cell to respond to the signal. For example, the spemann-mangold organiser can induce changes in the cells it affects only during a restricted time window. Once a cell has received an inductive signal it develops autonomously not receiving new signals from another cell. Therefore the response of a cell to a particular signal depends largely on the current state of that cell; an inductive signal can only select one response from a limited number of possible responses. This means that different signals can activate a particular gene at different stages of develoment turning the genes on and off when required. Also, the same signal can elicit different responses in different cells depending on their developmental history.
An example of induction can be seen in the development of the vertebrate eye in which the optic vesicle induces the overlying ectoderm to differentiate and form the lens placode. The lens then induces the overlying ectoderm to form the cornea.
Yet another example of induction can be illustrated with C.elegans. In formation of the vulva 3 of 6 precursors form the vulva (P3-P8) . P5-P7 form the vulva whilst the others form the epidermis. The anchor cell lies next to the P6 and secretes a diffusible signal inducing it to adopt the primary cell fate of the vulva. The next two nearest cells P5 and P7 receive a lower concentration and adopt a secondary fate thus forming the outer structure of the vulva. P6 then uses lateral inhibition to stop the remaining cells from becoming vulval tissue so they remain epidermal. (Sommer et al).
A summary of the process of induction:
- Induction may be permissive or inductive.
- It may be long-range or cell-to-cell.
- The same families of signalling molecules may elicit different responses in different cells depending on the developmental history of that cell.
- Inductive signals are transduced from the cell surface to the nucleus by intracellular signal transduction molecules (second messenger pathways).
- Within the nucleus transcription factors are activated and this results in changes in gene expression and therefore cell state.
As mentioned before cells may also become different from their neighbours through asymmetric cell division which is the process by which a cell divides to give two daughter cells with different fates due to the unequal segregation of cytoplasmic determinants.
Drosophila neuroblasts are stem cells that give rise to neuronal cells in the neuroepithelium of the neural plate. A single neuroblast delaminates (ingresses) from the neuroepithelium in the neuroectoderm. It then divides asymmetrically to give another neuroblast cell and a smaller ganglion mother cell (GMC). The GMC then divides to give two neuronal cells whilst the Neuroblast divides to give another Neurolast and GMC. During asymmetric division protein complexes form crescents at the apical and basal cortex of the Neuroblast. As the cell begins to divide in anaphase the chromatids separate via the spindle and the GMC inherits the basal proteins whilst the neuroblast inherits the apical proteins.
Another example of asymmetric division can be seen in C.elegans. P-granules segregate with the germline lineage as they are the cytoplasmic determinant of germ cell fate and are consequently packaged into P cells. Protein complexes are found in the anterior of the zygote anchored by the cytoskeleton network. The actin cytoskeleton is crucial as depolymerisation of actin filaments disrupts asymmetric cell division. In the anterior complex proteins such as Bazooka (par3), par6 and protein kinase C are found which are also found in the Drosophila neuroblasts. The position of the mitotic spindle is crucial. If the plane of division was not parallel to the segregated determinant then all of the daughter cells would inherit the same fate.
Another example of asymmetric cell division is the development of sensory organs in Drosophila. Drosophila have bristles which are a key sensory organ comprised of a hair cell, socket cell, sheath cell and a sensory neuron cell. In the sensory organ precursor (SOP) the ‘Numb‘ protein is localised at the cortex of the cell. Asymmetric division gives rise to Iia, without Numb, and Iib which inherits Numb. It functions by antagonising Notch signalling, a cell surface receptor. Notch binds to delta protein and together they affect cell fate. The Iia cell continues developing and later re-synthesises Numb protein so that both lineages Iia and Iib both express the Numb protein. In the Iia cell lineage, after a round of asymmetric division, the numb inherited cell becomes a hair cell whilst the other cell becomes a socket cell.
Lastly, another example of asymmetric cell division can be exemplified in the neurogenesis of the mammalian neocortex. Neurogenesis in the cerebral cortx of the mammalian brain occurs in the proliferative ventricular zone (PVC) – deepest layer. Symmetric and Asymmetric cell division occur. At mitosis, Numb and Notch are localised to opposite sides of the cell. When the spindle is parallel to the lumen both daughter cells inherit Numb and notch and Notch activity is repressed by Numb so they become undifferentiated stem cells. When the spindle is perpendicular to the lumen, one daughter cell inherits Numb protein and the other doesn’t. Notch signalling is blocked in the Numb inherited cell and it becomes a stem cell and stays at the lumenal surface. The other cell does not repress Notch signalling and it differentiates into a neural progenitor whilst migrating to the basal surface.
References
Brenner S: The genetics of Caenorhabditis elegans. Genetics 1974, 77:71-94.
Sulston JE, Horvitz HR: Post-embryonic cell lineages of the nematode, Caenorhabditis elegans. Dev Biol 1977, 56:110-156.
Sulston JE, Schierenberg E, White JG, Thomson JN: The embryonic cell lineage of the nematode Caenorhabditis elegans. Dev Biol 1983, 100:64-119.
Riddle DL, Blumenthal T, Meyer BJ, Priess JR (eds): C. elegans II. Cold Spring Harbor, NY: Cold Spring Harbor Laboratory Press; 1997.
The C. elegans Sequencing Consortium: Genome sequence of the nematode C. elegans: a platform for investigating biology. Science 1998, 282:2012-2018.
Fraser AG, Kamath RS, Zipperlen P, Martinez-Campos M, SohrmannM, Ahringer J: Functional genomic analysis of C. elegans chromosome Iby systematic RNA interference. Nature 2000, 408:325-330.